INTRODUÇÃOA inervação eferente da cóclea, também denominada inervação olivococlear, na maioria dos mamíferos, consiste basicamente de dois sistemas, o sistema eferente medial (SEM) e o sistema eferente lateral (SEL). O SEM origina-se de neurônios grandes localizados anterior, ventral e medialmente ao núcleo olivar superior medial, possui fibras mielinizadas que se projetam principalmente para a cóclea contralateral e fazem sinapse diretamente na base das células ciliadas externas (CCE).
O SEL origina-se em pequenos neurônios dentro e próximo ao núcleo olivar superior lateral, possui fibras não-mielinizadas que se projetam principalmente para a cóclea ipsilateral e fazem sinapse nas terminações aferentes sob as células ciliadas internas (CCI) (1-5). As fibras contralaterais do SEM cruzam a linha média, ao nível do assoalho do quarto ventrículo, unem-se às fibras ipsilaterais no núcleo vestibular e alcançam a periferia com o nervo vestibular. No meato acústico interno (MAI), através da anastomose de Oort (anastomose vestíbulo-coclear) atingem o órgão espiral (6).
Existe evidência de que as fibras eferentes acompanham o nervo vestibular inferior (4,7-10). O SEM e grande parte do SEL têm a acetilcolina como neurotransmissor principal, mas o SEL também pode usar o ácido gama-aminobutírico e outros neurotransmissores ou neuromoduladores, incluindo encefalinas e o peptídeo relacionado ao gene calcitonina (11). Apesar do profundo conhecimento anatômico e fisiológico do sistema eferente, muitas dúvidas ainda não foram esclarecidas sobre sua importância funcional no processamento auditivo.
Vários estudos experimentais são realizados na tentativa de elucidar dúvidas sobre seu verdadeiro papel. Um modelo experimental envolvendo a ausência deste sistema pode ser obtido através da secção de suas fibras. Em animais de laboratório, comumente a secção é realizada ao nível do assoalho do quarto ventrículo (12-15) ou ao nível do nervo vestibular inferior (16-19). Entretanto, baseado na anatomia descrita, o local mais adequado para completa eliminação do sistema eferente seria ao nível do nervo vestibular (6).
A secção ao nível do assoalho do quarto ventrículo, quando realizada na linha média, elimina apenas as fibras contralaterais do SEM, deixando as fibras ipsilaterais e o SEL.
A secção ao nível do nervo vestibular inferior pode ocorrer depois da anastomose vestíbulo-coclear, devido à dificuldade de visualização do MAI em animais de laborató- rio, deixando toda a inervação eferente coclear integra.
Além disso, esta seletividade exige uma maior retração do tronco encefálico, aumenta a chance de lesão do núcleo coclear e oferece mais risco ao nervo e artéria coclear que estão localizados abaixo do nervo vestibular inferior. O objetivo deste trabalho é descrever uma via de acesso ao nervo vestibular em chinchilas, baseado no acesso cirúrgico em cobaias (20), com descrição anatômica detalhada de seu osso temporal.
MATERIAL E MÉTODOAmostraOito Chinchillas laniger, saudáveis, do sexo masculino, adultas, pesando entre 500 e 750g foram anestesiadas com uma associação de cloridrato de cetamina (Ketalar®) e xylasina (Rompum®), via intramuscular, na proporção de 2:1 respectivamente, com um volume total de 1ml/kg (21).
Doses complementares de cloridrato de cetamina foram administradas conforme a necessidade para manuten ção de um plano profundo de anestesia. O animal foi mantido sobre um colchão térmico para preservação da temperatura retal em torno de 37,5oC, devido à facilidade de hipotermia.
CirurgiaO osso temporal da chinchila consiste anatomicamente em quatro regiões:
1) bula;
2) meato acústico externo e membrana timpânica;
3) cadeia ossicular e
4) labirinto.
A bula é dividida em região dorsal, mastoídea e labiríntica (Figura 1). A bula dorsal é facilmente palpável sendo a principal referência anatômica pré-operatória. Na face medial do osso temporal, observa-se a fossa para a paraflocula cerebelar, importante referência para acesso ao MAI, por estar lateral e cranialmente ao mesmo, e este com os foramens para os nervos: vestibular, facial e coclear (Figura 2).
O nervo vestibular está posicionado mais lateralmente; o nervo facial, medialmente e o nervo coclear, inferiormente (22). A cirurgia foi realizada com auxílio de um microscó- pio e apenas do lado direito de cada animal. Com bisturi lamina 11, a pele e tecido celular subcutâneo foram incisados, confeccionando um retalho quadrado, para exposi ção da bula dorsal bilateral do osso temporal e dos músculos occipitais.
A musculatura cervical foi desinserida do osso temporal lateralmente e do osso occipital medialmente, para visualização do osso occipital. Através de um perfurador de estapedotomia, a região do osso occipital a ser exposta foi demarcada, evitando-se desta forma uma grande fratura e conseqüentemente hemorragia por lesão do seio sigmóide.
Com a própria lâmina de bisturi, removeu-se o osso occipital delimitado, com exposi ção da dura-máter. A mesma foi incisada para possibilitar a aspiração da parte póstero-lateral do cerebelo e visualização da fossa para a paraflocula cerebelar e conseqüentemente do MAI com seus nervos. O extravasamento de um pouco de líquor nesta fase é comum, assim como sangramento durante a aspiração, porém controlável. O nervo vestibular foi então identificado e avulsionado com um ponteiro tipo baioneta com a extremidade curva (utilizado em cirurgia otológica). Nenhum dos animais morreu durante o procedimento cirúrgico.
Os animais foram então sacrificados com uma dose letal de pentobarbital de sódio (400mg/kg) e na craniotomia realizada verificou-se apenas a secção do nervo vestibular com integridade dos nervos facial e coclear (Figura 3). Este experimento está de acordo com as normas do COBEA (Colégio Brasileiro de Experimentação Animal) e do UFAW (The Universities Federation for Animal Welfare) e foi aprovado pela Comissão de Análise de Projetos de Pesquisa (CAPPesq) do Hospital das Clínicas da FMUSP (protocolo nº 646/00).
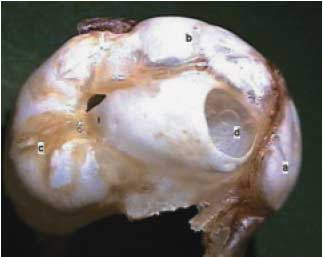
Figura 1. Visão lateral do osso temporal direito. (a) Bula dorsal, (b) bula mastóidea, (c) bula labiríntica, (d) meato acústico externo.
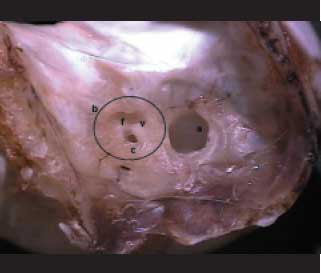
Figura 2. Visão medial do osso temporal direito. (a) Fossa para paraflocula cerebelar. Meato acústico interno (b) com os foramens para o nervo coclear (c), nervo facial (f) e nervo vestibular (v).
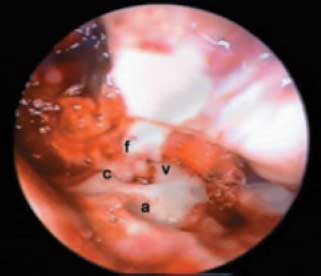
Figura 3. (a) Fossa para paraflocula cerebelar, (v) Coto distal do nervo vestibular, (f) nervo facial, (c) nervo coclear.
A secção do nervo vestibular em chinchilas foi um procedimento relativamente rápido e sem complicações. Como a secção foi realizada distal ao MAI, não foi necessário grande manipulação e retração do tronco encefálico, preservando desta forma as estruturas vizinhas.
Na cirurgia proposta por BARBARA et al. (1999) (20), em cobaias, é necessário a utilização do motor com broca de diamante para isolamento do seio sigmóide. Na nossa opinião, o uso do motor poderia influenciar as respostas das emissões otoacústicas caso fosse necessário sua realização no pósoperat ório, pois existe evidencia de que o ruído provocado pela broca leva a uma disfunção das CCE evidenciado pela alteração na amplitude das emissões otoacústicas por produtos de distorção em humanos (23).
Além disso, o isolamento do seio sigmóide poderia fazer com que a cirurgia se tornasse mais demorada e com risco aumentado de lesão vascular. Na via de acesso descrita, a osteotomia foi realizada abaixo do seio sigmóide, sem o risco de inadvertidamente, lesar algum seio venoso. Segundo BARBARA et al.(1999) (20), o nervo vestibular em cobaias está localizado mais lateralmente no MAI o que facilita a sua secção, e foi o que observamos também na chinchila. Apesar da secção das fibras eferentes ao nível do assoalho do quarto ventrículo ainda ser utilizada em trabalhos experimentais (14,15), ainda restam as fibras ipsilaterais do SEM e todo o SEL.
SCHUKNECHT (1959) (24) observa a persistência de acetilcolinesterase na região das CCE após a efetiva secção do SEM ao nível do assoalho do quarto ventrículo em gatos. O mesmo é observado em ratos por GODFREY e ROSS (1985) (25). ZHENG et al. (1999) (18) citam que poucas fibras eferentes remanescentes são suficientes para manter o funcionamento auditivo normal. Conseqüentemente, os resultados eletrofisiológicos da cóclea após a secção do SEM ao nível do assoalho do quarto ventrículo poderiam, ainda, estar sob influência da acetilcolina. ZHENG et al. (1997, 1999) observam que, mesmo após cuidadosa secção do nervo vestibular inferior em chinchilas, há a presença da acetilcolinesterase, verificada através da análise histológica da cóclea, em alguns animais (17,18).
Observa-se que no primeiro trabalho (17) de 8 orelhas submetidas a cirurgia, 5 apresentavam a acetilcolinesterase na região das CCE e CCI.
No segundo trabalho (18), 4 de 8 orelhas submetidas a cirurgia apresentavam a acetilcolinesterase na região das CCE e CCI. Estes dados sugerem que a cirurgia para secção do nervo vestibular inferior em chinchilas é extremamente difícil devido à dificuldade de visualização do MAI. A secção do nervo vestibular antes da sua entrada no MAI como fizemos, oferece várias vantagens:
a) Menor tempo cirúrgico;
b) O uso do motor com broca é desnecessário;
c) Não requer isolamento de seios venosos e
d) Menor manipulação de estruturas vizinhas.
Além disso, é utilizada em seres humanos para estudo do sistema eferente (26,27). O perfeito entendimento da técnica cirúrgica para acesso ao nervo vestibular é de grande importância para os experimentos na área da otologia, principalmente para o estudo do sistema eferente.
Com a descrição anatômica detalhada da face medial do osso temporal em chinchilas, e com a via de acesso proposta, objetivou-se facilitar os experimentos em laboratório quando for necessário o uso destes animais.
CONCLUSÃOA via de acesso descrita mostrou-se eficiente para a secção do nervo vestibular em chinchilas. O tempo cirúrgico foi relativamente rápido com pouca manipulação do tronco encefálico.
AGRADECIMENTOSAgradecemos ao Presidente da Associação Brasileira dos Criadores de Chinchila laniger, Sr. Carlos Luis Perez, pela colaboração em ceder os animais estudados neste trabalho.
REFERÊNCIAS BIBLIOGRÁFRICAS1. Warr WB. Olivocochlear and vestibular efferent neurons of the feline brain stem: their location, morphology and number determined by retrograde axonal transport and acetylcholinesterase histochemistry. J Comp Neurol, 161: 159-182, 1975.
2. Warr, WB, Guinan JJ. Efferent innervation of the organ of Corti: two separate systems. Brain Res, 173: 152-155, 1979.
3. Guinan, JJ, Warr, WB, Norris, BE. Differential olivocochlear projections from lateral versus medial zones of superior olivary complex. J Comp Neurol, 221: 358-370, 1983.
4. Liberman, MC, Brown, MC. Physiology and anatomy of single olivocochlear neurons in the cat. Hear Res, 24: 17-36, 1986.
5. Brown, MC. Morphology of labeled efferent fibers in the guinea pig cochlea. J Comp Neurol, 260: 605-618, 1987.
6. Spoendlin, H. Neural anatomy of the inner ear. In: Jahn, AF, Sacchi, JS. Phisiology of the ear, New York, Raven Press; 1988, p. 201-219.
7. Rasmussen, GL. The olivary peduncle and other fiber connections of the superior olivary complex. J Comp Neurol, 84: 141-219, 1946.
8. Gacek, RR. The efferent cochlear bundle in man. Arch Otolaryngol, 74: 690-694, 1961.
9. Ross, MD. The general visceral efferent component of the eighth cranial nerve. J Comp Neurol, 135: 453-478, 1969.
10. Littman, TA, Bobbin, RP, Cullen, JKJr. Histochemical evidence that cochlear efferents are carried with the inferior vestibular nerve in guinea pigs. Hear Res, 56: 281-285, 1991.
11. Guinan, JJ. Physiology of olivocochlear efferents. In: Dallos P, Popper AN, Fay RR, ed. The cochlea. New York, Springer-Verlag; 1996, p. 441-442.
12. Liberman, MC. Effects of chronic cochlear de-efferentation on auditory-nerve response. Hear Res, 49: 209-223, 1990. Murao MS Arq. Otorrinolaringol., São Paulo, v.9, n.2, p. 96-100, 2005.100
13. Puel, JL, Rebillard, G. Effect of contralateral sound stimulation on the distortion product 2F1-F2: evidence that the medial efferent system is involved. J Acoust Soc Am, 87: 1630-1635, 1990.
14. Kakigi, A, Hirakawa, H, Mount, RJ, Harrison, RV. The effects of crossed olivocochlear bundle section on transient evoked otoacoustic emissions. Hear Res, 110: 34-38, 1997.
15. Hsieh, YL, Chen, TJ, Lin, CH, Chen, SS. Effect of olivocochlear bundle lesion on locomotor activity in rats. Physiol Behav, 64: 179-184, 1998.
16. Littman, TA, Cullen, JKJr, Bobbin, RP. The effect olivocochlear bundle transection on tuning curves and acoustic distortion products. J Acoust Soc Am, 92: 1945-1952, 1992.
17. Zheng, XY, Henderson, D, Hu, BH, Ding, DL, McFadden, SL. The influence of the cochlear efferent system on chronic acoustic trauma. Hear Res, 107: 147-159, 1997.
18. Zheng XY, Henderson D, McFadden SL, Ding DL, Salvi RJ. Auditory nerve fiber responses following chronic cochlear de-efferentation. J Comp Neurol, 406: 72-86, 1999.
19. Zheng, XY, Mcfadden, SL, Henderson, D, Ding, DL, Burkard, R. Cochlear microphonics and otoacoustic emissions in chronically de-efferented chinchilla. Hear Res, 143: 14-22, 2000.
20. Barbara, M, Attanasio, G, Piccoli, F, Filipo, R. Vestibular neurectomy in the guinea-pig: a retrosigmoid approach. Acta Otolaryngol (Stockh), 119: 171-173, 1999.
21. Costa, DL, Erre, JP, Aran, JM. Aminoglycoside ototoxicity and the medial efferent system: I. Comparison of acute and chronic gentamicin treatments. Audiology, 37: 151-161,1998.
22. Browning, GG, Granich, MS. Surgical anatomy of the temporal bone in the chinchilla. Ann Otol Rhinol Laryngol, 87: 875-872, 1978.
23. Cruz, MJ, Fagan, P, Atlas, M, McNeill, C. Drill-induced hearing loss in the nonoperated ear. Otolaryngol Head Neck Surg, 117: 555-558, 1997.
24. Schuknecht, HG, Churchill, JA, Doran, R. The localization of acetylcholinesterase in the cochlea. Arch Otolaryngol, 69: 549-559, 1959.
25. Godfrey, DA, Ross, CD. Enzimes of acetylcholine metabolism in the rat cochlea. Ann Otol Rhinol Laryngol, 94: 409-414, 1985.
26. Scharf, B, Magnan, J, Chays, A. On the role of the olivocochlear bundle in hearing: 16 cases studies. Hear Res, 103: 101-122, 1997.
27. Scharf, B, Magnan, J, Collet, L, Ulmer, E, Chays, A. On the role of the olivocochlear bundle in hearing: a case study. Hear Res, 75: 11-26, 1994.
Trabalho realizado no Laboratório de Otorrinolaringologia da Faculdade de Medicina da Universidade de São Paulo . LIM 32, apresentado no American Academy of Otolaryngology - Head and Neck Surgery Foundation, em 19-22 de setembro de 2004.
Endereço para correspondência: Márcia Sayuri Murao . Rua Dona Cota, 177, sala 202 . Varginha / MG . CEP 37010-560. Telefone: (35) 3221-2730.
Fax: (35) 3212-7417. e-mail: marciamurao@hotmail.com
Artigo recebido em 30 de março de 2005. Artigo aceito em 14 de maio de 2005.
* Doutor em Otorrinolaringologia pela Faculdade de Medicina da Universidade de São Paulo.
** Professor Associado da Disciplina de Otorrinolaringologia da Faculdade de Medicina da Universidade de São Paulo.
*** Professor Assistente Doutor da Disciplina de Topografia Estrutural Humana da Faculdade de Medicina da Universidade de São Paulo.