INTRODUÇÃOEstudos que avaliam os efeitos da privação e da estimulação auditiva sobre os circuitos inibitórios (CHANG et al., 2003; KAPFER, 2002; KANDLER, 2004; RAJAN, 2002; SALVI et al., 2000; VALE et al., 2003) e sobre a percepção auditiva (BOÉCHAT, 2004) têm evidenciado de forma irrefutável que o desenvolvimento e o funcionamento do sistema auditivo estão relacionados à quantidade e à qualidade da entrada auditiva (input).
É clara a relação entre lesão coclear, geração de zumbido e intolerância a sons. Cerca de 78% a 90% dos pacientes com zumbido apresentam alteração nos limiares audiométricos (FUKUDA, 1990; MARTINS, 1991; COELHO et al., 2004). Aproximadamente 66% dos pacientes com zumbido têm algum grau de intolerância a sons e, desses, 33% buscam tratamento específico para a hiperacusia, e 86% dos pacientes com hiperacusia têm zumbido (JASTREBOFF e JASTREBOFF, 2004). Entre pacientes em busca de tratamento para hiperacusia, 53% apresentam também algum grau de perda de audição (JASTREBOFF e JASTREBOFF, 2004).
O objetivo deste artigo é revisar as descobertas recentes sobre os efeitos da privação auditiva nos circuitos inibitórios do SNAC e relacionar esses achados com estudos sobre zumbido e hiperacusia e com a prática clínica otorrinolaringológica e fonoaudiológica.
REVISÃO DA LITERATURAPara auxiliar o leitor na compreensão dos estudos revisados a respeito dos efeitos da privação auditiva nos circuitos inibitórios do SNAC, faremos uma breve revisão de conceitos básicos do funcionamento do SNAC (sugerimos leitura de ROMAND,1992; ROUILLER, 1992 e FERES, 1998).
O estudo da fisiologia de fibras individuais do SNAC mostra que cada fibra responde a uma faixa estreita e bem definida de freqüências. A análise das freqüências às quais uma única fibra responde e os limiares de cada uma das respostas é chamada de curva de sintonia da fibra. Nessa curva, o ponto de maior sensibilidade da fibra, ou seja, no qual a despolarização ocorre com menor limiar, é chamado de freqüência característica da fibra (FC). Partindo do limiar de estimulação da fibra nervosa, o número de potenciais de ação cresce de acordo com o aumento da intensidade do estímulo, até atingir um ponto máximo, chamado ponto de saturação. Após o ponto de saturação, o aumento na intensidade do estímulo não aumenta mais a freqüência de potenciais de ação. Chama-se domínio dinâmico da fibra a faixa de respostas que vai do limiar ao ponto de saturação. A freqüência característica, a atividade espontânea e o domínio dinâmico são propriedades que dão uma identidade única a cada fibra do nervo auditivo (ROUILLER, 1992).
Em cada núcleo da via auditiva central, as propriedades de respostas dos neurônios auditivos se modificam no sentido aferente, indicando que o sinal é processado de modo seqüencial e progressivo. Em quase todos os pontos da via auditiva há também projeções eferentes, o que sugere que os núcleos não são independentes; pelo contrário, estão sob controle de centros hierarquicamente superiores na via. Em outras palavras, o processamento da informação auditiva é possível graças à cooperação das diversas estações do sistema auditivo (ROUILLER, 1992).
Os sinais elétricos que carregam a informação acústica são transmitidos entre os neurônios por meio de sinapses. Na via auditiva, a grande maioria das sinapses é mediada por neurotransmissores (sinapses químicas), o que permite que o sinal seja modificado ao longo de seu trajeto e que, de acordo com a necessidade, haja mudanças plásticas no sistema. As sinapses mediadas pelo neurotrans¬missor glutamato, por exemplo, são chamadas de excita¬tórias, por aumentarem a possibilidade de que o neurônio pós-sináptico gere um potencial de ação, enquanto as mediadas por GABA e glicina, por exemplo (chamadas GABAérgicas e glicinérgicas), são consideradas inibitórias.
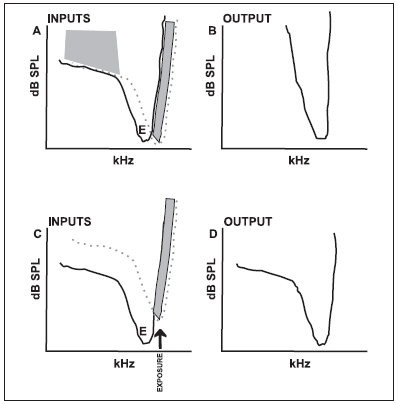
No sistema auditivo, assim como no visual e somatossensorial, os mecanismos de inibição aferentes partem da mesma área que provoca excitação no neurônio (RAJAN, 2001). A inibição ocorrerá quando a quantidade de informações inibitórias for maior que a de excitatórias no campo de recepção. Assim, nos pontos em que a inibição se sobrepõe às áreas de excitação da curva de sintonia da fibra auditiva, haverá um mascaramento dessas respostas excitatórias para determinadas freqüências (Figuras 1A e 1B).
Muito já se sabe sobre os circuitos excitatórios, que constituem aproximadamente 60% do SNC. Entretanto, os mecanismos que cercam a plasticidade e o desenvolvimento dos circuitos inibitórios do sistema nervoso central (SNC) ainda são pouco compreendidos (CHANG et al., 2003), pois estes são constituídos de uma rede neuronal morfologicamente e funcionalmente complexas. Na via auditiva, entretanto, os circuitos inibitórios são topograficamente bem organizados e precisos, servindo de modelo para o estudo da função inibitória (KANDLER, 2004). Ao contrário do que ocorre em mapas topográficos de outros sistemas sensoriais, que têm sinapses basicamente excitatórias (mediadas por glutamato e acetilcolina), a organização do mapa tonotópico do sistema auditivo tem diversas vias puramente inibitórias, mediadas por glicina, GABA ou ambas.
Figura 1. Ilustração esquemática do modelo de inibição por bandas. As figuras de cima mostram dados de uma orelha normal. As figuras de baixo são de uma orelha que sofreu trauma acústico. A: curva de sintonia mostrando a interação entre as entradas (input) excitatórias (linha sólida) e inibitórias (linha pontilhada) de um neurônio normal. A característica de freqüência (CF) da entrada inibitória está localizada em uma freqüência discretamente acima da FC da entrada excitatória. Um neurônio hipotético normal produz uma resposta excitatória (área aberta acima da curva de sintonia excitatória) desde que a entrada excitatória tenha um limiar mais baixo que o limiar de entrada inibitória. As respostas são inibidas, ou mascaradas (área sombreada), se a entrada inibitória tiver um limiar igual ou inferior ao da entrada excitatória. B: A saída (output) excitatória hipotética do neurônio normal em A. C: O mesmo modelo é mostrado, mas o limiar da entrada inibitória é elevado como resultado a uma exposição acústica (exposure) a um tom que traumatizou a freqüência acima da FC excitatória e próxima à FC inibitória. D: Igual a B, mas com saída do neurônio modificada pelo tom traumatizante. Note que a saída das respostas excitatórias são expandidas, principalmente nas áreas da curva que respondem às freqüências baixas. (Figura de SALVI et al., 2000, com permissão do autor).
As regiões mais estudadas da via auditiva que têm função inibitória são a oliva superior medial (que tem papel importante na localização sonora), a via entre núcleo medial do corpo trapezóide (NMCT) e a oliva superior lateral (OSL) (o NMCT envia informação puramente inibitória da orelha contra-lateral para OSL, que a integra com informações excitatórias da orelha ipsi-lateral) e o colículo inferior (maior centro de integração auditiva subcortical).
Pesquisas desenvolvidas nos últimos anos sugerem que os circuitos inibitórios desempenhem um importante papel na percepção de intensidade, na discriminação auditiva, no zumbido e na hiperacusia.
Evidências fisiológicas da plasticidade do SNAC por privação auditiva Plasticidade é um fenômeno biodinâmico que provoca mudanças de propriedades fisiológicas, bioquímicas e/ou anatômicas dos neurônios centrais em função da demanda de transmissão de informações acústicas. Assim, o sistema auditivo se reorganiza quando há uma variação na entrada auditiva (input), seja por diminuição da entrada, nos casos de lesões cocleares, ou por aumento, quando há uma nova entrada auditiva (durante o desenvolvimento pós-natal e após a colocação de implante coclear ou prótese auditiva, por exemplo).
O desenvolvimento e a manutenção de fibras do SNAC, assim como de outras estruturas do SNC, dependem de interações entre os neurônios pré e pós sinápticos. KAPFER et al. (2002) estudaram o desenvolvimento da primeira estrutura do SNAC responsável pela localização sonora de freqüências baixas no plano horizontal, a oliva medial superior (OMS) em cobaias. A OMS recebe estímulos excitatórios (glutamato) e inibitórios (glicina), das duas orelhas e compara a diferença de tempo interaural. Como essa diferença pode ser de apenas alguns microssegundos, as sinapses glicinérgicas precisam ser altamente precisas. Para otimizar a precisão temporal pós-sináptica as entradas glicinérgicas se dão próximos à zona de disparo do neurônio (corpo celular ou dendritos proximais). No entanto, esse refinamento só é atingido logo após duas semanas de vida com estimulação auditiva. Nas cobaias recém-nascidas, nas cobaias jovens com ablação coclear unilateral e nas que receberam estímulo sonoro unidirecional observou-se sinapses glicinérgicas estão uniformemente distribuídas no corpo celular, dendritos proximais e distais. Portanto, o desenvolvimento e a organização da rede neuronal inibitória, fundamental para a sintonia fina, é dependente de atividade (KAPFER et al., 2002).
Havendo perda de elementos pré-sinápticos, alteram-se eventos inter e intracelulares. Nos neurônios pós-sinápticos pode haver alterações de estrutura, função e metabolismo do mesmo, podendo, muitas vezes, levá-lo à morte neuronal (ZIRPEL et al., 1997). Desse modo, a lesão coclear pode causar mudanças dramáticas nas respostas excitatórias nos neurônios corticais que servem ao sistema auditivo.
Quando há uma lesão parcial da cóclea, regiões centrais que antes respondiam pelas regiões cocleares lesadas passam a ser ocupadas pelas entradas vindas de regiões preservadas da cóclea. Em outras palavras, os mapas tonotópicos de todo o SANC podem ser alterados pela expansão da representação de freqüências cujo limiar de percepção é normal ou melhor (WILLOT, 1996; RAJAN, 2001).
Outro efeito sofrido no SNAC após uma lesão coclear é o desmascaramento, ou seja, manifestação de inputs excitatórios que antes eram inibidos (RAJAN, 2001). Nesse caso, mesmo uma pequena lesão, permanente ou temporária no órgão receptor, ou uma condição que simule uma perda auditiva, podem levar à perda de inibição imediata ao redor de uma única célula do SNAC (Figuras 1C e 1D) (RAJAN, 2001; SALVI et al., 2000).
WANG et al. (1996) avaliaram as respostas do núcleo do colículo inferior e determinaram suas curvas de freqüências (tuning curve). Observaram fibras que só eram excitadas por uma faixa bem estreita de freqüências próximas a 8 kHz e cuja descarga aumentava até aproximadamente 20 dB acima do limiar e decaía consideravelmente para intensidades mais elevadas. Após a exposição da orelha contralateral a 16 kHz (uma oitava acima) a 107 dB NPS por 20 minutos o limiar da FC do neurônio e a resposta para freqüências altas não sofreram alterações. No entanto, o limiar para freqüências baixas aumentou até 25 dB nas freqüências de 4 a 5 kHz e a fibra passou a responder para freqüências bem mais baixas (500 Hz), para as quais antes não respondia. Tais mudanças ocorreram em cerca de 40% nos neurônios do colículo inferior (CI), ou seja, a maior parte das fibras do CI não foi afetada pela exposição traumática. O que diferencia as células afetadas das não afetadas são suas características funcionais, que sugerem que as últimas não possuam entradas inibitórias laterais importantes.
Em um estudo semelhante, SALVI et al. (1996) avaliaram as conseqüências da exposição a sons intensos no núcleo coclear dorsal (NCD). Os achados mostraram que os neurônios com zonas excitatórias estreitas (que respondem a poucas freqüências) e zonas inibitórias tiveram as mesmas mudanças funcionais encontradas no CI, enquanto as células com respostas amplas a freqüências tiveram mudanças mínimas.
Os efeitos da lesão de CCI por carboplatina sobre a atividade elétrica na janela redonda, no CI e no córtex auditivo primário foram pesquisados por meio de poten¬ciais auditivos evocados (PAE) nessas três regiões (WANG et al., 1997). Os dados revelaram diminuição de 50% dos PAE da janela redonda, pouca ou nenhuma alteração na função de entrada e saída do CI para intensidades fracas e moderadas e redução de até 50% para intensidades mais fortes, e aumento da amplitude dos potenciais do córtex auditivo, com grande variabilidade inter-animais. O pico da amplitude dos PAE no córtex auditivo (acima do normal) foi atingindo algumas semanas após a lesão e então se reduzindo um pouco. Os achados sugerem que exista um controle de ganho entre a cóclea e o tronco encefálico que compense ou reduza os efeitos de uma lesão de CCIs. Também parece haver um controle de ganho do córtex auditivo que visa ajustar o ganho auditivo para mantê-lo na faixa operacional normal (WANG et al., 1997; SALVI et al., 2000).
Evidências clínicas da plasticidade do SNAC por privação e estimulação auditiva Com o objetivo de comparar o desempenho na percepção de intensidade entre usuários de AASI e não usuários, PHILLIBERT et al. (2002) avaliaram 18 sujeitos com DANS bilateral simétrica (64-90 anos), sendo 9 não usuários de AASI e 9 usuários de AASI (pelo período de 1 a 5 anos) nas duas orelhas (digitais com controle de ganho), pareados por idade, gênero e grau de perda. Os autores aplicaram a Escala de Sensação de Intensidade (loudness scale) e o Limite de Discriminação de Intensidade (DLI ou Discimination-limen for intensity), que avalia a habilidade do indivíduo em detectar pequenas diferenças de intensidade. Os resultados mostraram diferenças significativas nos dois testes. Usuários de AASI percebem diferenças menores de intensidade e classificam como "ok" sons que para os não-usuários eram classificados como "forte". Assim, parece haver mudanças na codificação de intensidade de sujeitos usuários de AASI.
Já a sensação de intensidade do zumbido parece não ser alterada em função do tempo de instalação da perda de audição. OCHI et al. (2003) fizeram a medição da loudness e do pitch do zumbido de pacientes com deficiência auditiva unilateral e zumbido unilateral (N=132). Os sujeitos foram divididos de acordo com o tempo de duração dos sintomas. Um grupo foi formado por pacientes com zumbido e DA aguda (surdez súbita há menos de 7 dias) e o outro grupo composto por paciente com zumbido e deficiência auditiva crônicos (por mais de 3 meses, de origem idiopática). Os autores correlacionaram as medidas loudness e do pitch do zumbido ao tempo de instalação da perda auditiva. As diferenças estatisticamente significantes entre os dois grupos foram a média etária, menor para o grupo de surdez e zumbido súbitos (média de 44,0 anos ± 13,6, contra 54,0 anos ± 15,9 no grupo de DA e zumbido crônicos) e a distribuição do pitch do zumbido. A distribuição do pitch do zumbido do grupo de surdez e zumbido súbitos apresentou-se difusa, com média de 5,81 kHz (DP = 3,22 kHz), em freqüências um pouco abaixo daquelas onde se encontra a maior diferença de limiar audiométrico entre orelha afetada e a não-afetada. No grupo de DA e zumbido crônicos o pitch do zumbido foi mais alto, com média de 2,96 kHZ (DP = 3,30 kHZ) e com dois picos de distribuição de pitch do zumbido, uma um pouco abaixo da freqüência com maior diferença de limiar audiométrico entre orelha afetada e não-afetada e outro 1/3 de oitava acima do pitch do zumbido. Portanto, observou-se o pitch do zumbido correspondeu a 0,2 oitava abaixo da freqüência de maior assimetria auditiva entre as orelhas, mas que sua relação com o tempo de instalação da perda auditiva é diferente, o que sugere que o córtex auditivo dos pacientes com DA e zumbido crônicos tenha sofrido uma reorganização.
BOÉCHAT (2003) verificou a interferência do tempo de privação e tempo de estimulação na variação da sensibilidade auditiva para tons puros e índice percentual de reconhecimento de fala (IPRF). Avaliou 72 sujeitos (idades de 10 a 86 anos) com DANS assimétricas bilaterais ou unilaterais, sendo 43 usuários de AASI na pior orelha e 29 não usuários de AASI na pior orelha. Considerando três avaliações audiológicas (inicial, intermediária e final) foram analisadas as variações para sensibilidade de tom puro e IPRF entre os grupos no período máximo de 6 anos em relação ao tempo de estimulação, tempo de privação, grau de assimetria entre orelhas e grau de perda. Como o grupo usuário de AASI teve menor variação dos limiares tonais e melhores IPRF, a autora concluiu ter havido tanto uma plasticidade por privação auditiva nos não-usuários de AASI na pior orelha, como uma plasticidade secundária após estimulação auditiva nos usuários de AASI. Observou também que nos dois primeiros anos após a introdução do estímulo auditivo há um maior ganho no uso do AASI, enquanto os efeitos negativos da privação se apresentam gradualmente ao longo do tempo de maneira homogênea. Boéchat considerou que a quantidade e a qualidade de estímulo determinam o desenvolvimento e a manutenção do SNAC e ressaltou a importância da estimulação auditiva binaural.
Efeitos do silêncio na via auditiva normalGeralmente o zumbido é melhor percebido pelos pacientes quando esses estão em locais silenciosos e sem desenvolver atividades que demandem atenção. O exemplo mais comum dessa situação é quando o indivíduo se deita para dormir, pois o incômodo com a percepção do zumbido pode dificultar ou até impedir seu sono.
São raros os estudos que avaliaram o efeito do silêncio na via auditiva de indivíduos com audição normal. O estudo de HELLER e BERGMAN (1953) é lembrado até hoje, por ter demonstrado que 94% de sujeitos sem queixa de perda de audição (sem comprovação audiométrica) apresentaram zumbido após permanecerem cinco minutos em uma câmara silenciosa. Seguindo a mesma linha de pesquisa, TUCKER et al. (2005) avaliaram o efeito do silêncio no surgimento da percepção do zumbido em 120 adultos (60 homens e 60 mulheres) com idades entre 18 e 30 anos e com audição normal. Após permanecerem em uma cabina acústica por 20 minutos, os indivíduos foram questionados sobre quaisquer sons que tivessem escutado durante o período de silêncio. No total, 64% dos sujeitos (n=77) relataram ter percebido sons no silêncio (incluindo sons referentes à pulsação cardíaca), com diferença significativa entre raças (78% de caucasianos e 38% de negros perceberam o zumbido) e sem diferenças entre gêneros.
Se considerarmos que grande parte da população fica em ambientes relativamente silenciosos ao menos no momento de dormir, seria de se esperar, então, muito mais relatos de percepção de zumbido em normo-ouvintes do que realmente ocorre. Talvez a informação dada aos participantes das pesquisas citadas tenha ativado a atenção auditiva dos sujeitos pois, ainda que não houvesse a sugestão de que esse som pudesse vir de dentro deles mesmos, estes eram previamente informados de que poderiam ouvir alguns sons enquanto estavam dentro da câmara.
Tratamentos atuais sugerem que pacientes incomodados com o zumbido devem receber, além do tratamento para a(s) doença(s) de base, sons competitivos que diminuem o contraste entre o zumbido e o ambiente (JASTREBOFF e HAZELL, 1993; HAZELL, 2002; SANCHEZ, 2002; SANCHEZ e FERRARI, 2003; KNOBEL et al, 2004; JASTREBOFF e JASTREBOFF, 2004).
A Terapia de Habituação ou TRT (Tinnitus Retraining Therapy) foi desenvolvida no início da década de 1990 com o intuito de promover a habituação de reações induzidas pelo zumbido e pela intolerância a sons (JASTREBOFF e HAZELL, 1993). Envolve sessões individuais de orientação e acompanhamento e enriquecimento sonoro. O tipo de enriquecimento sonoro é indicado segundo as queixas auditivas do paciente (grau de incômodo com o zumbido, tipo de intolerância a sons e grau de perda auditiva, se presentes). Em linhas gerais, recomenda-se o uso constante de sons suaves, contínuos e monótonos para todos os tipos de paciente. Esses sons podem ser apresentados no ambiente (música, sons da natureza, fontes de água, ventilador, etc), diretamente no meato acústico (geradores de som pessoais) ou pela amplificação sonora de sons ambientais por meio de AASI, para os portadores de deficiência auditiva. Na TRT, a adaptação bilateral de aparelhos geradores de som ou dos AASI (se houver DA bilateral) deve ser sempre bilateral, mesmo quando o zumbido do paciente é unilateral. A permanência em locais silenciosos, assim como o uso de protetores auditivos em locais sem ruídos intensos, são contra-indicados (JASTREBOFF e HAZELL, 1993; JASTREBOFF e JASTREBOFF, 2004).
DISCUSSÃOFisiologicamente, o zumbido pode ser explicado por qualquer mudança na sincronia neural ou nos padrões de descarga elétrica da atividade neural espontânea das vias auditivas (EGGERMONT, 2003). O surgimento agudo do zumbido após uma lesão coclear pode ser explicado pela perda de inibição lateral, que desmascararia respostas excitatórias de circuitos neurais antes inibidos (SALVI, 2000; RAJAN, 2001). Assim, o desmascaramento de novas respostas excitatórias sugerem que a perda da inibição tem um papel permissivo crítico (RAJAN, 2001; OCHI et al, 2003). Já os zumbidos crônicos e as intolerâncias a sons podem estar relacionados a uma hiperatividade gradual do córtex auditivo como conseqüência de uma lesão coclear (SALVI, 2000) e à reorganização cortical (OCHI et al, 2003).
Os estudos aqui apresentados evidenciam mudanças plásticas nas porções da via auditiva que codificam a intensidade. Essas mudanças ocorrem em função da quantidade e a qualidade da estimulação acústica e se refletem na percepção de intensidade. Os achados de SALVI (2000) sugerem que exista um controle de ganho entre a cóclea e o tronco encefálico que compense ou reduza os efeitos de uma lesão de CCIs. Também parece haver um controle de ganho do córtex auditivo que visa ajustar o ganho auditivo para mantê-lo na faixa operacional normal. É provável que nos pacientes com hiperacusia esse controle de ganho não esteja operando normalmente, o que geraria a percepção anormal de sons fracos e moderados (JASTREBOFF e JASTREBOFF, 2004).
JASTREBOFF e JASTRBOFF (2004) propõem que a privação auditiva por diminuição da saída coclear (output) pode gerar zumbido ou exacerbar um zumbido pré-existente. Essa relação parece estar relacionada às mudanças plásticas que ocorrem no sistema nervoso auditivo central (SNAC) após a lesão do órgão receptor, e não às conseqüências diretas da lesão coclear (SALVI, 2000).
Os achados de BOÉCHAT (2003) atribuem um valor fundamental à protetização precoce e à estimulação equilibrada entre as orelhas e chega a sugerir que perdas auditivas bilaterais simétricas com adaptação de AASI monoaural podem se tornar assimétricas em virtude da estimulação assimétrica. Esses dados também dão suporte à recomendação da TRT de que o uso de geradores de som seja sempre bilateral e que se mantenha ao máximo a simetria entre as orelhas (JASTREBOFF e HAZELL, 1993; JASTREBOFF e JASTREBOFF, 2004). Em nossa prática clínica observamos que pacientes com zumbido que adaptam AASI ou gerador de som apenas na orelha acometida pelo zumbido (zumbido unilateral) ou na orelha com pior zumbido (zumbido bilateral) passam a perceber o zumbido na orelha contra-lateral. Acreditamos que, a curto-prazo, a melhora ou o mascaramento do zumbido na pior orelha facilite a percepção de um zumbido, antes subclínico, na outra orelha.
A exploração aprofundada dos princípios que regem a organização funcional das vias inibitórias do sistema auditivo pode proporcionar o entendimento de anormalidades na organização de circuitos inibitórios que parecem estar presentes no zumbido e na intolerância a sons. A partir de então, novas estratégicas terapêuticas poderão ser desenvolvidas.
COMENTÁRIOS FINAISA literatura tem progressivamente fornecido evidências de plasticidade das vias auditivas que sugerem que a prática diária otorrinolaringológica e fonoaudiológica deve ser revisada no que diz respeito a situações atualmente pouco valorizadas. Assim, alertamos para os benefícios do diagnóstico precoce das situações de privação auditiva, zumbido e intolerância a sons, bem como para o seu tratamento, seja pela protetização auditiva precoce e bilateral nos casos de perda auditiva, ou para o uso de estimulação sonora bilateral e simétrica nos casos de zumbido e intolerância a sons.
REFERÊNCIAS BIBLIOGRÁFICAS1. Boéchat EM. Plasticidade do sistema auditivo quanto à sensibilidade auditiva para tons puros e respostas para a fala na deficiência auditiva neurossensorial. São Paulo, 2003,175p. (Tese de Doutorado - Faculdade de Medicina da USP).
2. Chang EH, Kotak VC, Sanes DH. Long-term depression of synaptic inhibition is expressed postsynaptically in the developing auditory system. J Neurophysiol, 2003, 90(3):1479-88.
3. Dauman R, Bouscau-Faure, F. Assessment and amelioration of hyperacusis in tinnitus patients. Acta Oto-Laryngologica, 2005, 125: 503-509.
4. Féres MCLC. Estudo das respostas plásticas de núcleos cocleares secundárias à deafarentação unilateral por ablação de cóclea em ratos Wistar neo-natos. [tese - doutorado - Universidade Estadual de São Paulo]. Ribeirão Preto, 1998.
5. Hazell JWP, Sheldrake JB, Graham RL. Decreased sound tolerance: predisposing factors, triggers and outcomes after TRT. In: International Tinnitus Seminar, 7. 2002. Fremantle, Australia. Proceedings of VII International Tinnitus Seminar, Perth, Australia: The University of Western Australia, 2002. p.255-61.
6. Jastreboff P, Jastreboff MM. Tinnitus Retraining Therapy (TRT) as a Method for Treatment of Tinnitus and Hypercausis Patients. J. Am. Acad. Audiol., 2000, v.11, n.3, p. 162-177.
7. Jastreboff P, Jastreboff MM. Decreased Sound Tolerance. In: SNOW J. Tinnitus: Theory and Management. Hamilton: BC Decker, 2004. p. 8-15.
8. Kandler K. Activity-dependent organization of inhibitory circuits: lessons from the auditory system. Curr Opin Neurobiol, 2004, 14: 96-104.
9. Kapfer C, Seidl AH, Schweizer H, Grothe B. Experience-dependent refinement of inhibitory inputs to auditory coincidence-detector neurons. Nat Neurosci, 2002, 5:247-253.
10. Ochi K, Ohasi T, Kenmochi M. Hearing Impairment and tinnitus pitch in patients with unilateral tinnitus: comparison of sudden hearing loss and chronic tinnitus. Laryngoscope, 2003, 113: 427-431.
11. Phillibert B, Collet L, Vesson J-F, Veuillet E. Intensity-related performances are modified by long-term hearing aid use: a functional plasticity? Hear Research, 2002, 165: 142-151.
12. Rajan R. Plasticity and inhibition in the receptive field of primary auditory cortical neurons after limited receptor organ damage. Cerebral Cortex, 2001, 11:171-182.
13. Romand R. Aspects anatomo-fonctionnels du noyau cochléaire. In: Romand R. (Ed). Le systeme auditif central. Anatomie et physiologie. Les Éditions Inserm, Paris, 1992, 51-108.
14. Rouiller EM. Organisation fonctionnelle des voies auditives. In : Romand R. Le Systeme Auditif Central. Les Éditions Inserm, Paris, 1992, 1-49.
15. Salvi RJ, Wang J, Ding D. Auditory plasticity and hyperactivity following cochlear damage. Hear Research, 2000, 147: 261-274.
16. Tucker DA, Phillips SL, Ruth RA, Clayton WA, Royster E, Todd AD. The effect of silence on tinnitus perception. Otolaryngol Head Neck Surg, 2005, 132(1):20-4.
17. Vale C, Schoorlemmer J, Sanes DH. Deafness disrupts chloride transporter function and inhibitory synaptic transmission. J Neurosci, 2003, 23:7516-7524.
18. Wang J, Salvi RJ, Powers N, Plasticity of response properties of inferior colliculus neurons following acute cochlear damage. J Neurophysiol, 1996, 75: 171-183.
19. Wang J, Powers NL, Hofstetter P, Trautwein P, Ding D, Salvi R. E¡ects of selective inner hair cell loss on auditory nerve fiber threshold, tuning and spontaneous and driven discharge rate. Hear. Res., 1997, 107:67-82.
20. Willot JF. Physiological plasticity in the auditory system and its possible relevance to hearing aid use, deprivation and acclimatization. Ear Hear, 1996, 17:66-77.
21. Zirpel L, Lachica EA, Rubel EW. Afferent regulations of neurons in the avian cochlear nucleus: intracellular mechanisms and signal transduction pathways. In: Berlin C. (ed). Neurotransmission and hearing loss - Basic science, diagnosos and management. San Diego: Singular Publing Group, 1997, p. 77-89.